本文
海藻類に関する研究
ノリの優良種苗開発試験
近年、海水温上昇の影響により秋季の水温降下時期が遅れ、ノリ養殖の開始時期が遅れる傾向にあります。また、ノリ養殖の開始後にノリ葉体に高水温による障害(葉体の変形や脱落)が発生し、生産に影響を与えています。
そこで、愛知県漁業協同組合連合会と共同で高水温被害を軽減できるスサビノリ種苗としてあゆち黒誉れとあゆち黒吉を開発し、種苗法に基づく品種登録出願を行いました(あゆち黒誉れは出願中、あゆち黒吉は平成24年4月登録、平成28年4月登録期間満了)。あゆち黒誉れは高水温での障害が少なく葉体の色調が濃いことが特徴で、あゆち黒吉は高水温でも単胞子(*)の放出量が多く葉体の色調が濃いことが特徴です。この2品種は混合して養殖することによって、生産されるノリ製品の品質向上と生産量の増大をはかることができ、使用している県内の生産者からは高評価を得ており、さらなる普及を図っています。
なお、あゆち黒誉れは海外への持ち出しが制限されています。
県内のノリ養殖を行っている海域では、さらなる高水温化や貧栄養化などが引き続き問題となっており、今後もこうした環境の変化の影響を削減できる優れた形質をもつ種苗の開発を目指します。
*単胞子とはノリの葉体から遊離した細胞のことで、ノリ網に付着して新たな葉体として生長します。


高水温(24℃)で2週間培養したあゆち黒誉れ(上)と標準品種U-51(下)の比較

養殖したあゆち黒吉葉体(上)と単胞子を放出するあゆち黒吉葉体(下)
バリカン症対策技術の開発
西三河地区の河口域漁場では、近年、ノリ生産量が大幅に減少しており、原因の一つには柵当たりの生産枚数が減少傾向にあることがあげられます。生産性が悪化している主な原因として、近年、被害が増えている養殖ノリのバリカン症があげられます。バリカン症は全国的にみられるノリの障害で、発生すると養殖されているノリ葉体の多くが途中で切れてしまいます。調査の結果、矢作川河口域でのバリカン症の発生と張り込み水位や塩分濃度は関係はなく、ヒドリガモ等の食害生物による食害が原因である可能性が高いと考えられました。今後は、その他の原因生物についてや、より効果的な対策について研究を行う必要があります。

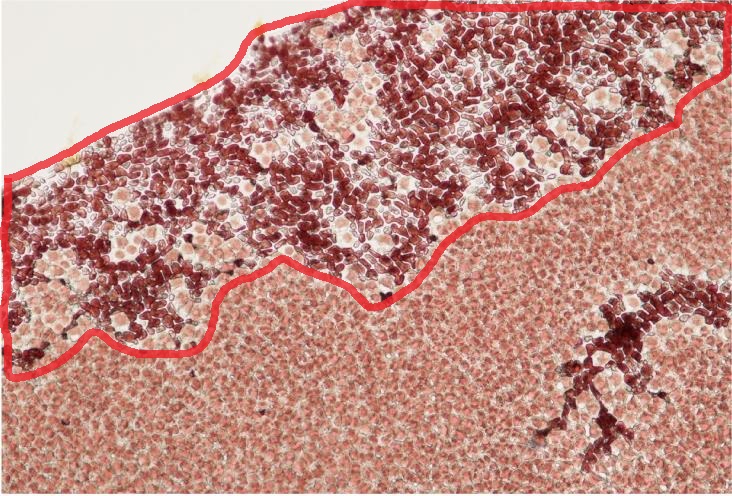
西尾漁場においてバリカン症を発症したノリ養殖網(左)とノリ葉体(右)
ノリの病障害に関する研究
スミノリ症はノリの病気のひとつで、本県では、年末年始にかけてしばしば発生して被害を出しています。スミノリ症のノリの細胞は、淡水に浸漬すると細胞の中身が飛び出してしまうため、ノリを真水で漉く工程で、製品が艶のない色調となって品質が低下してしまいます。症状が重いと製品にすることすら困難になります。被害を大きくしないためには、病気の兆候をできるだけ早期に見つける必要があります。
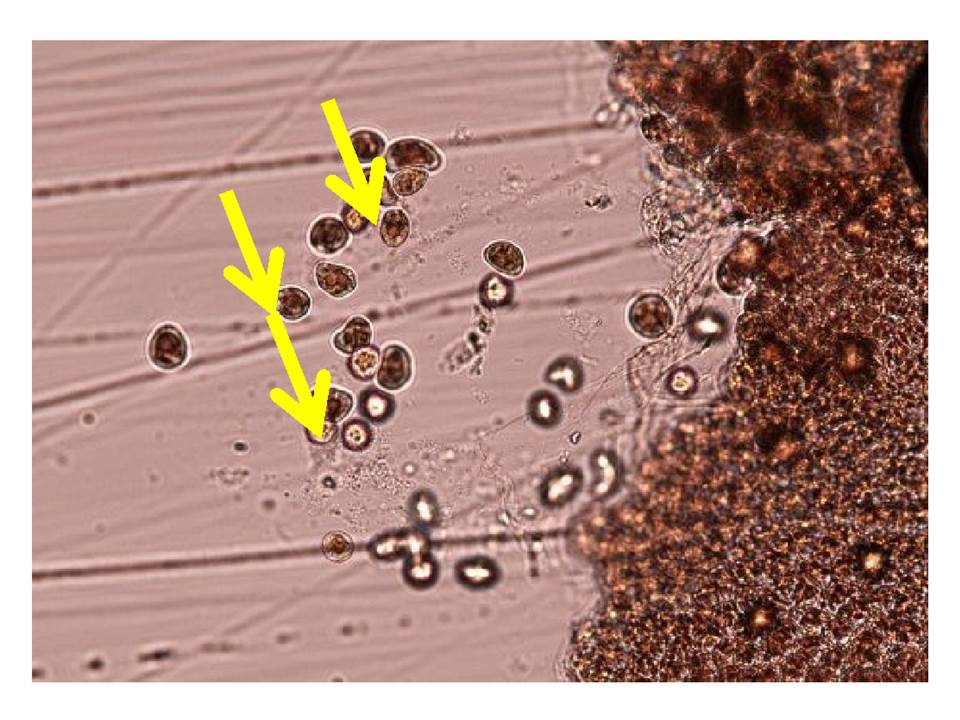
淡水に浸漬したスミノリ症のノリ葉体。赤枠部で細胞の中身が飛び出している。

正常な製品とスミノリ症のノリ葉体からつくられた製品。右2枚の製品は艶がない。

スミノリ症検査の泳動像 赤枠部のバンドが陽性を示す。
参考文献
※1 三宅佳亮・植村宗彦・伏屋 満(2005)愛知県内ノリ養殖漁場から分離されたスミノリ症原因菌のPCRによる検出.愛知水試研報,11,17-24.
※2 伏屋 満・ニノ方圭介・植村宗彦・盛田 信(2001)室内培養における結合型残留塩素およびFlabobacterium sp.による養殖ノリのスミノリ症の発症.愛知水試研報,8,15-20.
サガラメ・カジメの藻場を再生する技術の開発
伊勢・三河湾の湾口に広がる沿岸岩礁域では、大型多年生コンブ目褐藻のサガラメとカジメ(以下、カジメ類)が藻場を成しています。これらはアワビ類の餌となり、メバルやイカ類の生育・産卵場となるなど、沿岸漁業にとり重要な役割を果たしています。

愛知県沿岸岩礁域のサガラメ

愛知県沿岸岩礁域のカジメ
しかし平成10年以降、サガラメ・カジメ群落は衰退し、未だ回復していません。
その原因は、海水温の上昇で、カジメ類に生理的に悪影響を及ぼしたこと(※1)と、湾口部で多数確認されている植食性魚類のアイゴ(※2)の摂食が活発になり摂食期間も長くなったこと(※1)が考えられます。


愛知県沿岸のおけるサガラメの凋落
また、カジメ類の藻場が衰退した海域では、ウニ類やアメフラシが認められます。これら底生の植食動物により、わずかな幼い葉体(胞子体)も摂食されてしまうため、藻場が再生できないと考えられます。さらに、母藻が減ったため種(遊走子)が不足している可能性があります。
そこで藻場を再生するため、これまでに、アルギン酸ナトリウムを用いたサガラメ幼胞子体の移植法(※3)や、サガラメ側葉の基部に生分解性繊維を絡ませアイゴの食害から生長点を保護する技術(※4)などを開発しました。

アルギン酸ナトリウムを用いたサガラメ幼胞子体の移植
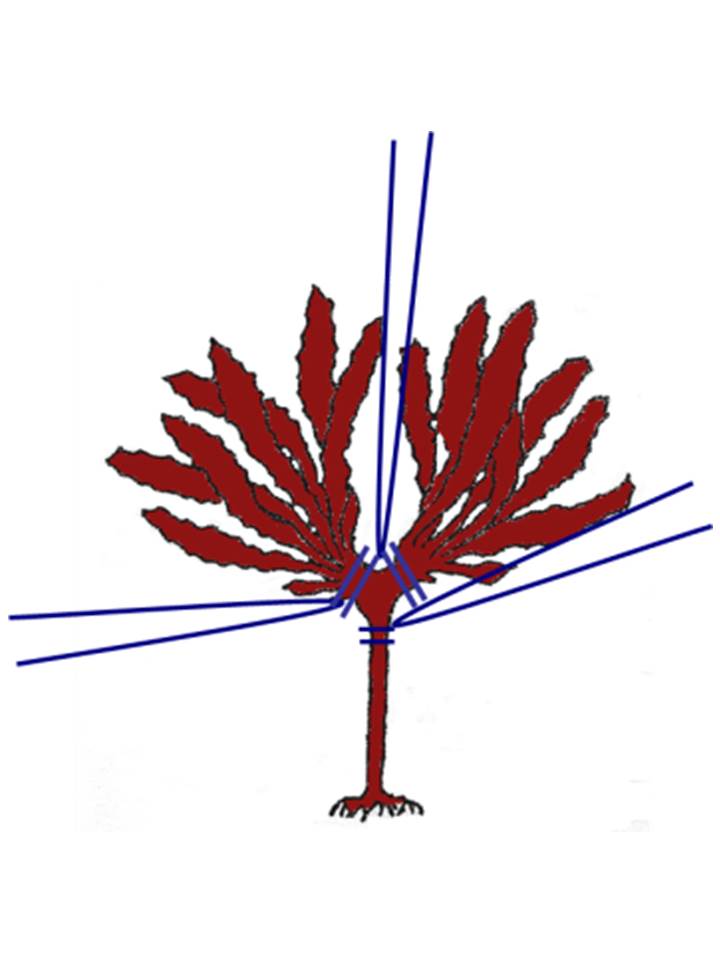

生分解性繊維を絡ませたサガラメ生長点の保護技術
平成22~24年には、これら技術を発展させるため、三重大学大学院、サカイオーベックス(株)、TBR(株)、及び(株)シャトー海洋調査との共同研究(※5)に取り組みました。そして以下の成果により,効率的に増殖させた配偶体または幼胞子体を、環境負荷の少ない素材を使って、既存の基盤に移植する低コストの藻場再生手法を開発しました。
1 サガラメ配偶体の成熟を制御する物質を明らかにし(※6)、大量培養装置を開発して培養効率を4倍向上させました。
2 包帯や組紐(※7,8)がサガラメの移植基質に利用できることを把握しました。
3 トゲモクとサガラメをノリ養殖用鋼管に混生移植することを可能にしました。
4 サガラメ母藻からの遊走子拡散範囲を求め、1平方メートル当たり1個体のサガラメ群落を1ha再生するために800個体の母藻が必要であることを把握しました。
平成30年に実施した藻場の調査では、藻場の再生技術の開発に取り組んだ南知多町(豊浜)の沿岸では、サガラメ藻場が大きく広がる様子を確認できています。


包帯や組紐を用いて移植した配偶体及び幼胞子体から生長したサガラメ
参考文献
※1 阿知波英明・落合真哉・芝 修一(2014):愛知県沿岸におけるサガラメ・カジメ分布面積の変動と衰退要因.愛知水試研報,19,38-43.
※2 蒲原 聡・伏屋 満・原田靖子・服部克也(2007):1997年から2005年までの愛知県岩礁域におけるサガラメEisenia arborea群落の様相.愛知水試研報,13,13-18.
※3 蒲原 聡・伏屋 満・柳澤豊重・服部克也(2007):アルギン酸ナトリウムと砂の混合ゾルに混入させたサガラメ幼葉の海底基質への移植法.水産工学,43(3),201-207.
※4 蒲原 聡・佐藤嘉洋・原田靖子・服部克也・鈴木輝明・高倍昭洋(2009):サガラメEisenia arboreaの分裂組織を生分解性繊維で保護する簡便なアイゴ採食防御法.水産工学,46(2),127-134.
※5 農林水産省農林水産技術会議の「平成22~24年度新たな農林水産政策を推進する実用技術開発事業(課題名:既存着底基質への海藻種苗の移植による効率的な藻場再生技術の実証試験)」
※6 倉島 彰・山本清春(2012):ジベレリンとステビオサイドによるコンブ目3種配偶体の成熟と生長の制御.藻類,60(1),9-14.
※7 伏屋 満・阿知波英明・落合真哉(2013):伸縮包帯とネット包帯を用いたサガラメとカジメのノリ養殖用鋼管柵への移植.愛知水試研報,18,33-34.
※8 阿知波英明・伏屋 満・青山 勧・山下 修(2013):組紐及び包帯を移植基質として用いた褐藻サガラメの生長.愛知水試研報,18,35-36.
問合せ
愛知県 水産試験場 漁業生産研究所
電話: 0569-65-0611
ファックス: 0569-65-2358
Adobe Readerをお持ちでない方は、バナーのリンク先からダウンロードしてください。(無料)
